From land management through habitat heterogeneity to biodiversity in grassland ecosystems: A combined theoretical, experimental, and observational study
In the first phase of the project, we want to focus on hypotheses 1-6 (see below). In the next phase, we plan to test predictions derived from hypothesis 7 as well as new predictions derived from the theoretical and empirical findings of the first phase.

Temperate grassland communities harbor a great fraction of the Central European plant species diversity and responses of these communities to anthropogenic disturbances have been investigated by numerous studies during the last decades. Our research focuses on one aspect that has been overlooked in previous studies: the role of micro-scale spatial heterogeneity in habitat conditions. This question is planned to be investigated in two phases. In the present phase we use a series of microcosm experiments and related models to uncover the fundamental mechanisms by which spatial heterogeneity in habitat conditions (habitat heterogeneity) affects the diversity of grassland communities. Our models and experiments are based on a novel concept termed ‘the area-heterogeneity trade-off’. The results obtained from this phase will be used as a baseline for planning a second phase of experiments that will be designed to test predictions obtained from our models and microcosm experiments under natural field conditions. Importantly, although our research focuses on grassland communities, we expect that our theoretical and empirical findings would provide general insights that will be applicable to studies focusing on larger scales and other kinds of organisms.
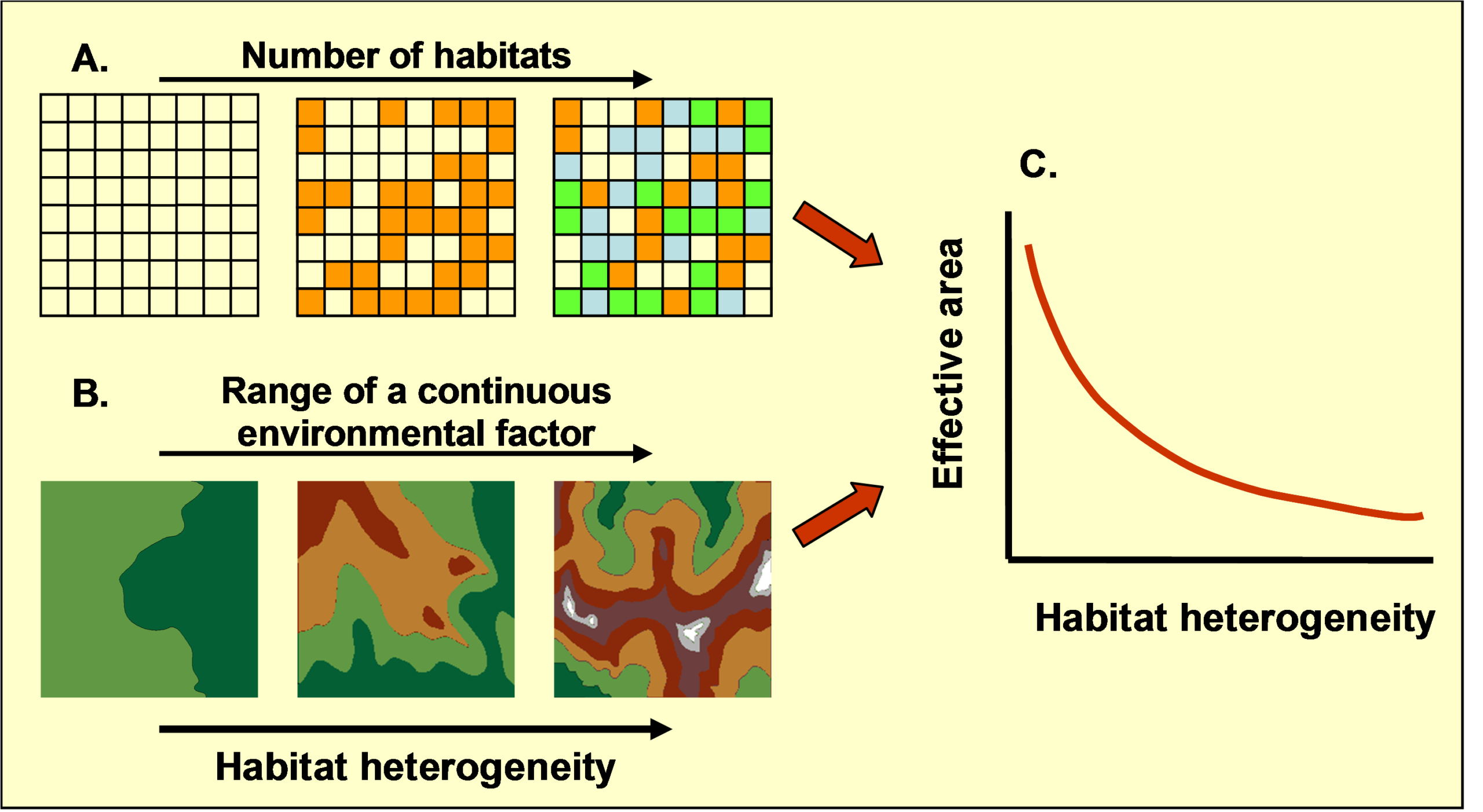
The ‘area-heterogeneity trade-off’ (Allouche et al. 2012) states that any increase in habitat heterogeneity must be associated with a corresponding decrease in the average amount of suitable area available per species (Fig.1). This geometric trade-off leads to a number of testable hypotheses:
1. Increasing habitat heterogeneity should increase the potential number of species in a community by providing suitable conditions for a larger number of species with different ecological requirements (this is the traditional prediction of niche theory).
2. At the same time, increasing heterogeneity reduces average population size (due to the proposed area-heterogeneity trade-off).
3. The reduction in population size should increase the likelihood of stochastic extinction.
4. The increase in extinction rates should reduce the number of species.
5. These contrasting positive and negative effects should lead to unimodal heterogeneity-diversity relationship, with habitat availability limiting richness at low levels of heterogeneity and area limiting richness at high levels of heterogeneity.
6. Specialist species should be more sensitive to the area-heterogeneity trade-off than generalist species and should therefore show stronger responses (both positive and negative) to variation in habitat heterogeneity.
7. The position of the inflection point (i.e., the level of heterogeneity that maximizes species diversity) should depend on properties of the relevant system. In general, any factor that increases the likelihood of stochastic extinction (habitat fragmentation, environmental stress, disturbance) should shift the inflection point to lower levels of heterogeneity and vice-versa.
The latter hypothesis implies that natural grassland communities can be expected to show various responses (positive, negative, unimodal, or flat) to increased habitat heterogeneity, depending on properties of the relevant system.
The core of the study is a system of microcosm experiments in which artificial communities of herbaceous plants are grown under different levels of ‘habitat heterogeneity’ (Fig. 2). The composition of the artificial communities was determined to represent species that occur in all three exploratories. The treatments used as sources of heterogeneity represent four factors that are known to be important in structuring grasslands of the exploratories (and temperate grasslands in general): soil depth, grazing/mowing (mimicked by clipping), fertilization, and trampling. The experiment includes two levels of soil depths, two levels of fertilization (with/without), two levels of clipping (with/without), and two levels of trampling (with/without). Together, these treatments result in 16 types of distinct ‘habitats’ and are applied using a factorial design to produce five levels of habitat heterogeneity (Fig. 2). By monitoring the response of the artificial communities to these manipulations we will be able to test whether and how population sizes, extinction rates, and species richness of the component species are influenced by micro-scale habitat heterogeneity.
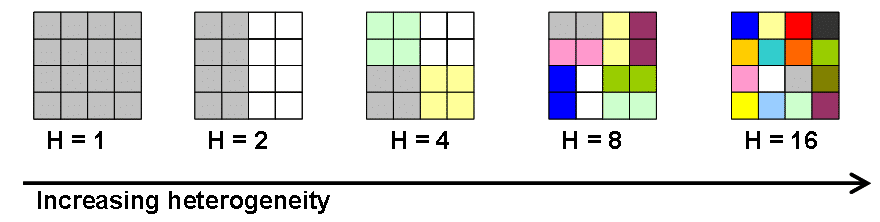
A microcosm approach enables us to avoid confounding factors that may bias the results of similar experiments conducted under field conditions. A previous study has indicated that such approach is highly effective as a means for testing hypotheses concerning mechanisms of species diversity in herbaceous plants (Ben-Hur et al. 2012). Though field experiments are planned to be conducted in a second phase of the research, we believe that microcosm experiments would serve as a better starting point for testing our hypotheses.
The modeling part of the project will provide a theoretical framework for the whole project and will rely on the area-heterogeneity trade-off as a unified concept. This trade-off was derived from a general modeling framework termed the Markovian Community Dynamics (MCD) framework (Allouche & Kadmon 2009) which is highly flexible in the kind of mechanisms that can be taken into account. In the proposed project we will use the MCD framework to derive analytical solutions that relax a number of simplifying assumptions that were made in the original formulation of the concept (e.g., a single source habitat for each species, fully random dispersal). We will also develop a spatially explicit framework for modeling the effects of continuous environmental variation, distance-limited dispersal, and spatial autocorrelation in habitat conditions, on the predictions of the model. We believe that the integration of these analytical and numerical approaches will allow us to significantly improve our understanding of the mechanisms by which fundamental properties of the species and the environment affect the ecological consequences of the area-heterogeneity trade-off.
In the current phase of the project we plan to focus on hypotheses 1-6 (see above). In the next phase we plan to test predictions derived from hypothesis 7, as well as new predictions that will be derived from the theoretical and empirical findings of the first phase.
Public Datasets
Project in other funding periods

Scientific assistants





